| |
|
深度读帖后再提问,在线时间过短不予回复。
所有信息仅供参考! |
|
| |
 都说免疫不入脑,入了就不得了
免疫治疗以后的疗效反应,当病灶增大时,不一定是进展,还有一种可能,叫做CR,这个现
都说免疫不入脑,入了就不得了
免疫治疗以后的疗效反应,当病灶增大时,不一定是进展,还有一种可能,叫做CR,这个现
 晚期肺癌经过4次换药,如今母亲已经4
作者:pears
“万事开头难”这句话在母亲的抗癌路上体现得淋漓尽致,最初确诊肺癌后半
晚期肺癌经过4次换药,如今母亲已经4
作者:pears
“万事开头难”这句话在母亲的抗癌路上体现得淋漓尽致,最初确诊肺癌后半
 既要,又要,还要,什么药品可以治疗
作者:seacat
EGFR突变可以分为常见突变和罕见突变。常见突变就是EGFR外显子19del突变
既要,又要,还要,什么药品可以治疗
作者:seacat
EGFR突变可以分为常见突变和罕见突变。常见突变就是EGFR外显子19del突变
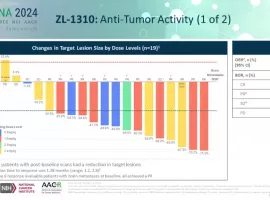 小细胞肺癌新希望?标准治疗耐药后OR
作者:Keenman
如果以药物规模化获批进入临床的时间来划分的话,进入21世纪以来,肺癌
小细胞肺癌新希望?标准治疗耐药后OR
作者:Keenman
如果以药物规模化获批进入临床的时间来划分的话,进入21世纪以来,肺癌
 母亲的基因检测和PD-L1报告出来了,
看了基因检测报告,似乎没有合适的靶向药,母亲确诊肺腺癌晚期,求助大家帮忙分析一下
母亲的基因检测和PD-L1报告出来了,
看了基因检测报告,似乎没有合适的靶向药,母亲确诊肺腺癌晚期,求助大家帮忙分析一下






 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡

